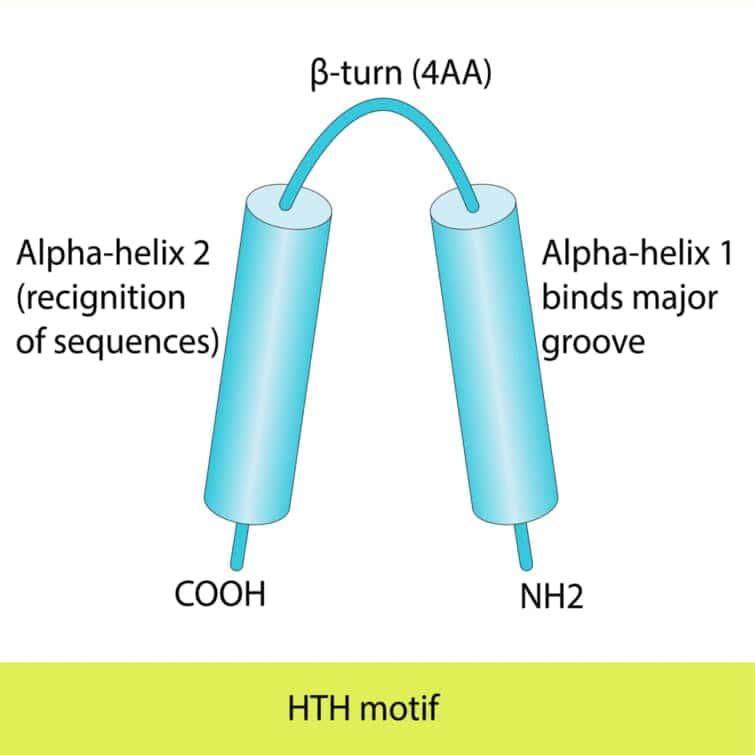
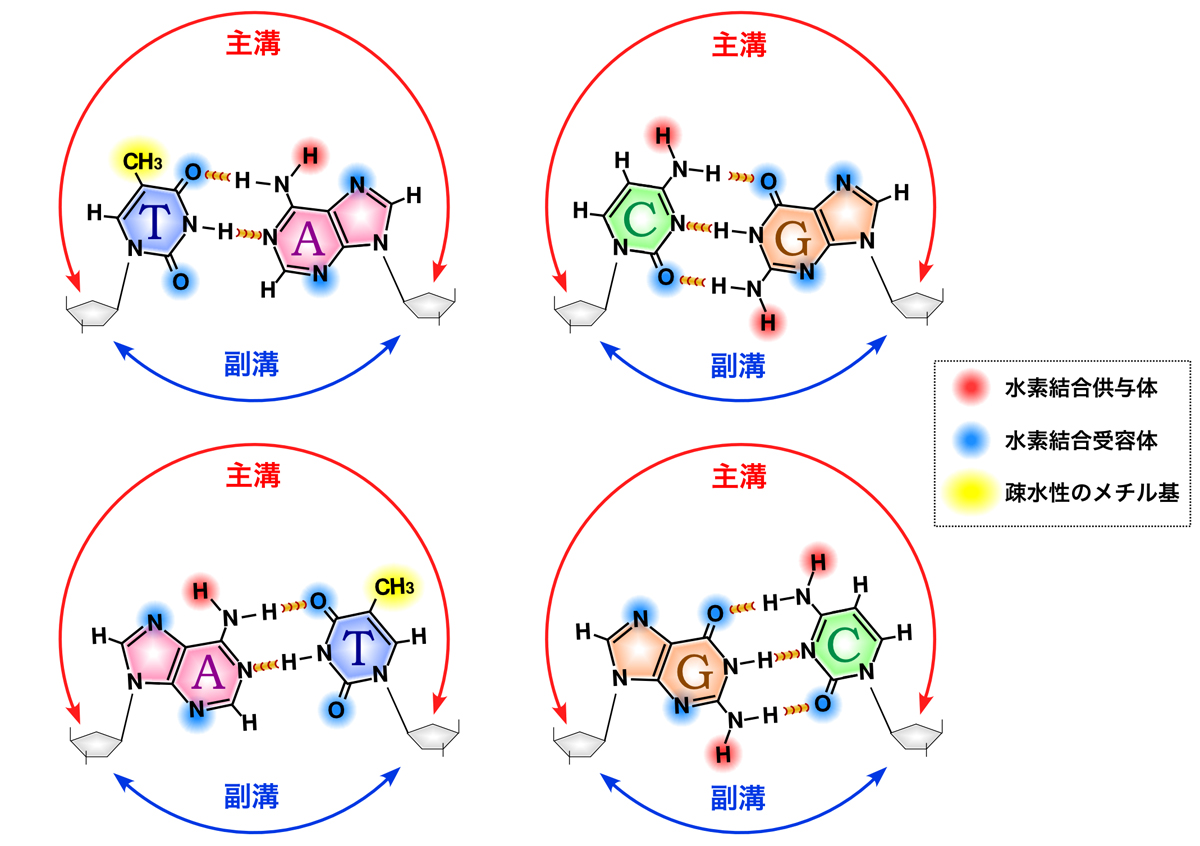
DNA結合ドメイン (DNA-binding domain: DBD)とは、二本鎖または一本鎖DNAを認識する少なくとも1つの構造モチーフを構造に含むタンパク質ドメインである。DBDは特定のDNA配列(認識配列)に対して特異的な、またはDNA一般に対して親和性を持つ。一部のDNA結合ドメインは、折り畳まれた構造の核酸も含む。
機能
DNA結合ドメインは、多くの場合、異なる機能を備えた別のタンパク質ドメインと共に大きなタンパク質を構成する。この他のドメインは、しばしばDNA結合ドメインの活性を調節する。DNA結合ドメインの機能は構造の形成、もしくは転写制御への関与、あるいはこの両方である。
DNA構造に関与するDNA結合ドメインはDNA複製、修復、保存、およびメチル化などのDNA修飾を担う。
遺伝子発現の調節に関与する多くのタンパク質はDNA結合ドメインを含む。例えば、転写因子と呼ばれるタンパク質は、DNAとの結合によって転写を調節する。ほとんどの細胞シグナル伝達カスケードの最終段階は、DNA結合タンパク質が関わる遺伝子調節である。
DNA結合ドメインは、DNA配列特異的または非特異的にDNAのヌクレオチドと相互作用するが、非特異的であってもタンパク質とDNA間の相互作用には何らかの分子相補性がある。DNA結合ドメインは、DNAの主溝または副溝で、あるいはDNAの構成糖とリン酸によってDNAを認識する。配列特異的なDNA認識は、タンパク質の機能や目的に合わせられている。例えば、デオキシリボヌクレアーゼの一種であるDNAse IはDNAをほぼランダムに切断するため非特異的である必要があるものの、DNAの特定の立体構造を認識し、DNAフットプリントと呼ばれる手法によるDNA認識の研究に有用な、ある程度特異的なDNA切断パターンを示す。
多くのDNA結合ドメインでは、特定の遺伝子を活性化する転写因子や、 制限酵素やテロメラーゼような特定の部位でDNAを修飾する酵素など、DNA配列を特異的に認識する必要がある。
DNA結合タンパク質の特異性は、ゲル電気泳動、分析的超遠心分離 、 熱量測定、DNA突然変異、タンパク質構造の突然変異または修飾、 核磁気共鳴 、 X線結晶構造解析、 表面プラズモン共鳴 、 電子スピン共鳴、架橋およびマイクロスケール熱泳動 (MST)など、多くの生化学的および生物物理学的手法を使用して研究されている。
ゲノムのDNA結合タンパク質
各ゲノムの大部分の遺伝子は、DNA結合タンパク質をエンコードしている(下表を参照)。ただし、DNA結合性を有するのは少数のタンパク質ファミリーのみである。例えば、約750個のジンクフィンガータンパク質を含む、約20,000個のヒトタンパク質のうち2000個程度のみがDNA結合性である。
種類
ヘリックスターンヘリックス
もともと真正細菌で発見されたヘリックス・ターン・ヘリックスモチーフはリプレッサータンパク質によく見られ、約20アミノ酸である。真核生物では、ホメオドメインは2つのヘリックスを含み、そのうちの1つはDNAを認識する認識ヘリックスである。これは、発達過程を調節するタンパク質によく見られる。
ジンクフィンガー
ジンクフィンガードメインは主に真核生物で見られ、一部は細菌でも発見されている。ジンクフィンガードメインは一般的に23〜28アミノ酸長であり、規則的な間隔で配置された残基(ヒスチジンまたはシステイン)で亜鉛イオンに配位結合することにより安定化し保持する。最も一般的なジンクフィンガー(Cys2His2)は、単一の亜鉛イオンを安定化し、1個の認識ヘリックスと1個のβシートで構成されており、1個の亜鉛イオンを安定化させる。
ロイシンジッパー
塩基性のロイシンジッパー ( bZIP )ドメインは主に真核生物で見られ、一部は細菌で見いだされる。bZIPドメインでは、7残基ごとに、ロイシンを含むαヘリックスが配置されている。あるヘリックスが他のヘリックスを見つけると、このヘリックスのロイシンはジッパーの歯のように相互作用し、2つのタンパク質を二量体化することを可能にする。DNAに結合すると、塩基性アミノ酸残基が糖リン酸骨格に結合し、ヘリックスが主溝に入り込み、遺伝子発現を調節する。
ウィングドヘリックス
ウィングドヘリックス (winged helix:WH)ドメインは、約110個のアミノ酸から成り、4個のヘリックスと1個の2ストランド2ストランドβシートを持つ。
ウィングドヘリックスターンヘリックス
ウィングドヘリックスターンヘリックス (winged helix-turn-helix:wHTH)ドメインは、通常85〜90アミノ酸長であり、1個の3ヘリックス束と4ストランドβシート(ウィング)によって形成されている。
ヘリックスループヘリックス
塩基性ヘリックスループヘリックス (basic helix-loop-helix:bHLH)ドメインは、いくつかの転写因子において見られる、2個のαヘリックスが1個のループで繋がっている構造をしている。通常、一方のヘリックスは他方よりも小さく、ループの柔軟性により他方のヘリックスに対して折り畳まれて包み込まれることにより、二量体化がなされている。通常、DNA結合領域は大きなヘリックスにある。
HMGボックス
HMGボックスドメインは、DNA複製や転写などの多くのDNA依存プロセスに関与する高移動度群タンパク質に見られる。また、屈曲を誘発することによりDNAの柔軟性を変化させる 。このドメインは、ループで区切られた3つのαヘリックスで構成される。
Wor3ドメイン
Candida albicansのWhite-Opaque Regulator 3(Wor3)にちなんで名付けられたWor3ドメインは、それ以前に発見されたほとんどのDNA結合ドメインよりも進化の歴史上近い時代に出現し、その存在は少数の菌類に限定されている。
OBフォールドドメイン
OBフォールドとは、オリゴヌクレオチド/オリゴ糖結合性(oligonucleotide/oligosaccharide binding)に因んで名付けられた小さな構造モチーフである。OBフォールドドメインは70〜150アミノ酸残基である。OBフォールドは一本鎖DNAに結合するため、OBフォールドドメインで構成されたタンパク質は一本鎖DNA結合タンパク質である。
OBフォールドタンパク質は、DNA複製、DNA組換え、DNA修復、転写、翻訳、低温ショック応答、およびテロメアの維持に重要であることが判明している。
免疫グロブリンフォールド
免疫グロブリンドメインは、大きな複数のループで接続された1個のβシートを持ち、このβシートはDNAの主溝または抗原のいずれかを認識するのに役立つ。通常、免疫グロブリンタンパク質に見られるが、サイトカイン経路のシグナル伝達性転写因子にも存在する。これは、サイトカイン経路が進化の歴史上比較的新しいもので、独自の生化学システムが構築されたのではなく、既存のシステムを利用したためと推測されている。
B3ドメイン
B3 DNA結合ドメインは、制限エンドヌクレアーゼのEcoRIIとBfiI、及び高等植物由来の転写因子でのみ発見されており、通常100-120残基から成る。7個のベータシートと2個のアルファヘリックスが含まれており、DNA結合性擬似バレル構造を形成する。
TALエフェクター
TALエフェクターは、キサントモナス属の植物病原細菌に見られ、細菌の毒性や増殖を促進するために、宿主植物の遺伝子発現の調節に関与する。複数の33-35残基のタンデム反復配列のある中央領域が1つあり、この領域は、TALエフェクター結合部位にある単一のDNA塩基をコードする 。このタンデム反復配列内で、DNA塩基と直接相互作用するのは残基13のみでありこれが配列特異性を決定するが、他の残基はDNA骨格と相互作用してDNAとの結合を安定化する。各反復配列は一対のαヘリックスの形をとるが、反復配列で構成されたアレイ全体は右巻きのスーパーヘリックスを形成し、DNA二重らせんを包み込む。このアレイはDNA結合時に収縮することが示されており、TALエフェクターの反復配列アレイにある唯一のN末端からDNAのチミン認識を始めてDNAの収縮を行うというDNA認識メカニズムが提案されている。植物病原細菌のRalstonia solanacearum、真菌内部共生菌Burkholderia rhizoxinica および未同定の海洋微生物2種でTALエフェクター関連タンパク質が発見されている。これらのグループ間において、反復配列アレイの構造とDNA結合領域の配列情報は保存されており、総じてTALE様タンパク質と呼ばれている。
ガイドRNAのための利用
Streptococcus pyogenesのCRISPR/Casシステムは、標的DNA部位との塩基対相補性を備えたガイドRNAの生物工学的操作により、真核生物の野生および人工プロモーターの活性化と抑制の両方を可能にする。Cas9は、ヌクレアーゼをRNAガイドで標的DNAに誘導させるための標的DNA上の結合プラットフォームとして使用できる。ドメインCas9は、標的の調節ドメイン(活性化、抑制、またはエピジェネティックエフェクターなど)、またはゲノムエンジニアリングの汎用ツールであるエンドヌクレアーゼドメインで利用でき、その後に異なるガイドRNAで複数の遺伝子座をターゲットにできる 。その後、異なるガイドRNAを使用して複数の遺伝子座をターゲットにします。
脚注
外部リンク
- 転写因子のDNA結合ドメインのデータベース、 “DBD: a transcription factor prediction database”. Nucleic Acids Research 34 (Database issue): D74-81. (January 2006). doi:10.1093/nar/gkj131. PMC 1347493. PMID 16381970. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1347493/. “DBD: a transcription factor prediction database”. Nucleic Acids Research 34 (Database issue): D74-81. (January 2006). doi:10.1093/nar/gkj131. PMC 1347493. PMID 16381970. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1347493/. “DBD: a transcription factor prediction database”. Nucleic Acids Research 34 (Database issue): D74-81. (January 2006). doi:10.1093/nar/gkj131. PMC 1347493. PMID 16381970. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1347493/. “DBD: a transcription factor prediction database”. Nucleic Acids Research 34 (Database issue): D74-81. (January 2006). doi:10.1093/nar/gkj131. PMC 1347493. PMID 16381970. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1347493/. “DBD: a transcription factor prediction database”. Nucleic Acids Research 34 (Database issue): D74-81. (January 2006). doi:10.1093/nar/gkj131. PMC 1347493. PMID 16381970. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1347493/. 精製DNA結合ドメインを使用した転写因子の予測用データベース
- DNA結合モチーフの一覧表
- DNA Footprinting - MeSH・アメリカ国立医学図書館・生命科学用語シソーラス(英語)
- DNA-Binding Proteins - MeSH・アメリカ国立医学図書館・生命科学用語シソーラス(英語)
- PROSITEの DNA結合ドメイン